.png)
Basic Dimension
http://sexualreligion.blogspot.com/
Number Archive

Disclaimer:
- Genetics and genealogy are serious professions with a lot of hidden rules which can change the calculations considerably.
http://sexualreligion.blogspot.com/
Number Archive

OPTICALLY REVISED:
- August 2020:
Disclaimer:
- Genetics and genealogy are serious professions with a lot of hidden rules which can change the calculations considerably.
Chromosomes:
http://abri.une.edu.au/online/pages/inbreeding_coefficient_help.htm

http://abri.une.edu.au/online/pages/inbreeding_coefficient_help.htm
Explanation of inbreeding in animal populations relies on a few basic genetic principles. Genetic information is stored in Chromosomes. Chromosomes are made up of DNA. Genes are sections of DNA and occur in pairs. A particular gene will occur at a particular site (locus, plural is loci) in the DNA of a particular Chromosome. The different forms of a gene (usually 2) that can occur at that locus are called alleles. Where both alleles are the same at the locus, they are called homozygous. Where the alleles are different, they are called heterozygous. In general, the two alleles will have an equal influence on the performance of an animal. That is, the heterozygous form (both alleles are present) tends to have performance midway between the two homozygous forms. In a few cases, one allele will have the main (dominant) effect on an animal, while the other allele will only have an effect in its homozygous form. These are called dominant and recessive genes.




The Coefficient of Inbreeding:
The coefficient of inbreeding (as proposed by Sewell Wright in 1922) is the probability that two alleles at a randomly chosen locus are identical by descent. Note that alleles may be identical for other reasons, but the inbreeding coefficient (F) is just looking at the mathematical probability that the alleles have come from a common ancestor.

The Coefficient of Inbreeding (as proposed by Sewell Wright in 1922) is the probability that two alleles at a randomly chosen locus are identical by descent. Note that alleles may be identical for other reasons, but the inbreeding coefficient is just looking at the mathematical probability that the alleles have come from a common ancestor.
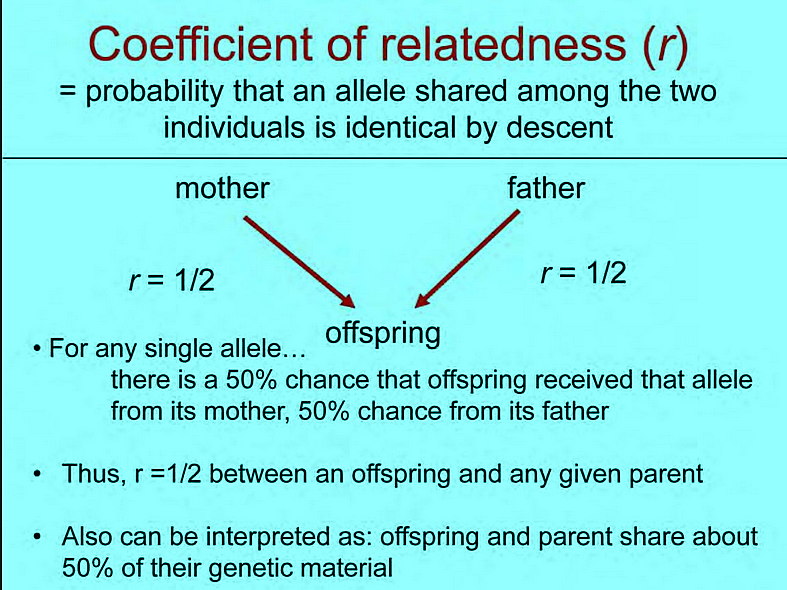
The Coefficient of Relationship (R) looks more a descriptive statistic of consanguinity relations within families. For example a father and his child have 50% of their genes in common. The same for two siblings. So it is not directly a probability for specific individual situations.
The Inbreeding Coefficient:

An important rule is the difference between independent chances (multiplication: ½x½=¼ [(2^-1)*(2^-1)=(2^-2)] and dependent chances (the addition of main effects supplied with multiplication for covariance).
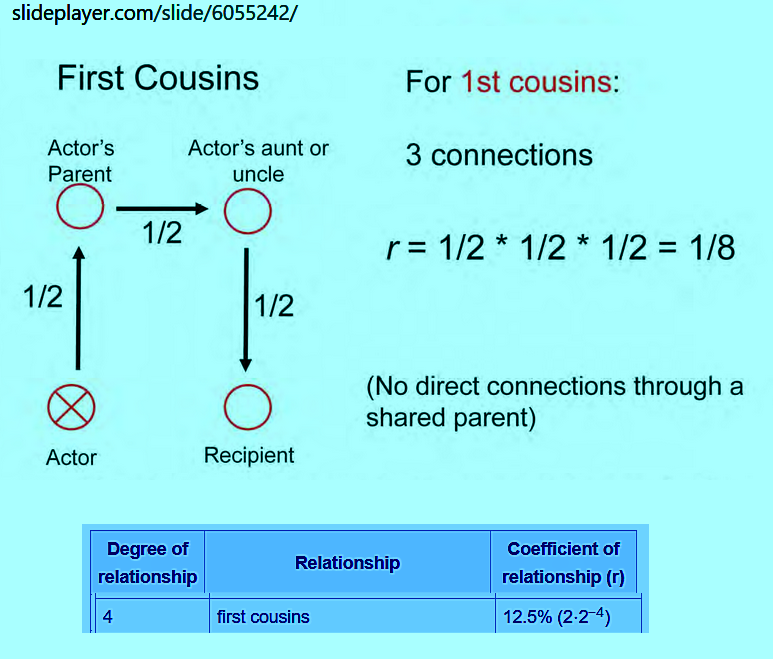
Other strategy first cousins:
Take, for example, the mating of first cousins who, by definition, share
a set of grandparents. For any particular gene in the male, the chance
that his female first cousin inherited the same gene from the same source
is 1/8. [AB x (BC or BD)=.5^3]. Further, for any gene the man passes to his child,
the chance is 1/8 that the woman has the same gene and ½ that she transmits
that gene to the child so 1/8 x ½ = 1/16. Thus, a first-cousin marriage has a
coefficient of inbreeding F =1/16= 6.25%.
F (coefficient of inbreeding): F is the symbol for the coefficient of inbreeding, a way of gauging how close two people are genetically to one another. The coefficient of inbreeding, F, is the probability that a person with two identical genes received both genes from one ancestor.
Autosomal disorders


https://www.youtube.com/watch?v=A-OU0JcuX6U
https://en.wikipedia.org/wiki/Coefficient_of_relationship
The coefficient of relationship (R) is a measure of the degree of consanguinity (or biological relationship) between two individuals. A coefficient of inbreeding (F) can be calculated for an individual, as a measure for the amount of pedigree collapse within that individual's genealogy. The term coefficient of relationship was defined by Sewall Wright in 1922, and was derived from his definition of the coefficient of inbreeding of 1921. The measure is most commonly used in genetics and genealogy.

[The more inbreeding (F) in the family the more the pedigree shrinks, collapses into a simpler model. In the limit of identical twins, people are equal, like Adam and Eve who at least had different sex chromosomes. Their pedigree crunched almost totally. Identical twins cannot breed, but Adam and Eve could, that's the power of their myth, which proves male sexual fantasy as an insane lust to inbreeding.]

The Coefficient of Relationship (R) looks more a descriptive statistic of consanguinity relations within families. For example a father and his child have 50% of their genes in common. The same for two siblings. So it is not directly a probability for specific individual situations:


Below we see the effect of both ancestors A and B on sibling C (green). We see the product rule of independent chances (of A and B) on an unspecified allele in sibling C:



Well, in statistics we can be flexible, it's no religion:





A common ancestor is any preceding individual in the pedigree who passes genes directly to two or more persons through different independent routes or paths:
http://www.genetic-genealogy.co.uk/supp/DirectRelationships.html http://www.genetic-genealogy.co.uk/Toc115570136.html
http://www.genetic-genealogy.co.uk/Toc115570135.html

http://www.genetic-genealogy.co.uk/supp/DirectRelationships.html

There are fewer kinds of direct relationship than collaterals and they are usually less complicated. This is because there are no half or first degree forms, which are a common feature of all collaterals. Furthermore, all direct relationships are intergenerational while collaterals can be members of the same or different generations. Finally with direct relationships, gene transmission is forward and unidirectional, in contrast to collaterals whose connections run in both directions of time through common ancestors.
A further fundamental difference is that inbreeding plays a more active role in the formation of double and multiple direct relationships. Most double and multiple collaterals are possible without inbreeding, but with direct relationships all plural forms result from matings between fairly close relatives.
Collateral relatives are related through common ancestors. They hold a proportion of their genes in common because they have received them from a common source. The important point is that no one receives any genes from collateral relatives.


Other strategy first cousins:
Take, for example, the mating of first cousins who, by definition, share
a set of grandparents. For any particular gene in the male, the chance
that his female first cousin inherited the same gene from the same source
is 1/8. [AB x (BC or BD)=.5^3]. Further, for any gene the man passes to his child,
the chance is 1/8 that the woman has the same gene and ½ that she transmits
that gene to the child so 1/8 x ½ = 1/16. Thus, a first-cousin marriage has a
coefficient of inbreeding F =1/16= 6.25%.
http://www.genetic-genealogy.co.uk/supp/DirectRelationships.html http://www.genetic-genealogy.co.uk/Toc115570136.html
http://www.genetic-genealogy.co.uk/Toc115570135.html







.jpg)

This work is licenced under a Creative Commons Attribution-Non Commercial-ShareAlike 4.0 International Licence.
https://en.wikipedia.org/wiki/Coefficient_of_relationship
The coefficient of relationship (R) is a measure of the degree of consanguinity (or biological relationship) between two individuals. A coefficient of inbreeding (F) can be calculated for an individual, as a measure for the amount of pedigree collapse within that individual's genealogy. The term coefficient of relationship was defined by Sewall Wright in 1922, and was derived from his definition of the coefficient of inbreeding of 1921. The measure is most commonly used in genetics and genealogy.
[The more inbreeding (F) in the family the more the pedigree shrinks, collapses into a simpler model. In the limit of identical twins, people are equal, like Adam and Eve who at least had different sex chromosomes. Their pedigree crunched almost totally. Identical twins cannot breed, but Adam and Eve could, that's the power of their myth, which proves male sexual fantasy as an insane lust to inbreeding.]
Coeff. of Inbreeding versus Coeff. of Relatedness:
The Coefficient of Inbreeding (as proposed by Sewell Wright in 1922) is the probability that two alleles at a randomly chosen locus are identical by descent. Note that alleles may be identical for other reasons, but the inbreeding coefficient is just looking at the mathematical probability that the alleles have come from a common ancestor.
The Coefficient of Relationship (R) looks more a descriptive statistic of consanguinity relations within families. For example a father and his child have 50% of their genes in common. The same for two siblings. So it is not directly a probability for specific individual situations.



Below we see the effect of both ancestors A and B on sibling C (green). We see the product rule of independent chances (of A and B) on an unspecified allele in sibling C:






A common ancestor is any preceding individual in the pedigree who passes genes directly to two or more persons through different independent routes or paths:
http://www.genetic-genealogy.co.uk/supp/DirectRelationships.html http://www.genetic-genealogy.co.uk/Toc115570136.html
http://www.genetic-genealogy.co.uk/Toc115570135.html
CALCULATION OF THE COEFFICIENT OF RELATIONSHIP
Common Ancestors Defined
Common ancestors are usually only shared by collateral [not interbreeding] relatives, since with direct [breeding] relationships one individual is the ancestor of the other. Exceptions are shown here.
In its literal sense a 'common ancestor' is any person who passes genes directly to two or more individuals. However, for the purposes of calculating the coefficient of relationship (R) and the coefficient of inbreeding (F), it has a more restricted definition:
In its narrow sense a common ancestor is any preceding individual in the pedigree who passes genes directly to two or more persons through different independent routes or paths. In the following diagram there are three individuals who satisfy this definition: i.e. B, H and I, who are common ancestors of J and K. (Note that parents are common ancestors in this context):
In its narrow sense a common ancestor is any preceding individual in the pedigree who passes genes directly to two or more persons through different independent routes or paths. In the following diagram there are three individuals who satisfy this definition: i.e. B, H and I, who are common ancestors of J and K. (Note that parents are common ancestors in this context):

http://www.genetic-genealogy.co.uk/supp/DirectRelationships.html
DIRECT RELATIONSHIPS
In male kin bonded tribes, genes are inherited by the male line systematically. But direct relations exist only between successive generations. Also in every generation there is an influx of new genes from fresh juvenile females from other tribes, which are a counterweight against inbreeding, these are also called direct relations but only for the immediate offspring:

There are fewer kinds of direct relationship than collaterals and they are usually less complicated. This is because there are no half or first degree forms, which are a common feature of all collaterals. Furthermore, all direct relationships are intergenerational while collaterals can be members of the same or different generations. Finally with direct relationships, gene transmission is forward and unidirectional, in contrast to collaterals whose connections run in both directions of time through common ancestors.
A further fundamental difference is that inbreeding plays a more active role in the formation of double and multiple direct relationships. Most double and multiple collaterals are possible without inbreeding, but with direct relationships all plural forms result from matings between fairly close relatives.
Collateral relatives are related through common ancestors. They hold a proportion of their genes in common because they have received them from a common source. The important point is that no one receives any genes from collateral relatives.
Collateral Relationship
An important rule must be observed for collaterals:
a) The path must go back against the arrows from one collateral relative to the common ancestor, and then follow the arrows forward to the other collateral relative. For example, in the next pedigree diagram, one of the paths between G and H is:
G_1_D_2_A_3_E_4_H
G_1_D_2_A_3_E_4_H
In this case trace the path back from one cousin to the common ancestor and then forward to the other cousin. Remember, a qualifying common ancestor is someone who passes genes directly to both collateral relatives along different routes. With collateral relatives there is often more than one common ancestor.
Find RGH
Common Ancestors [3]
|
Paths
|
(1/2)n
| ||
A
|
G_1_D_2_A_3_E_4_H
|
(1/2)4
|
=
|
1/16
|
B
|
G_1_D_2_B_3_E_4_H
|
(1/2)4
|
=
|
1/16
|
Σ(1/2)n
|
=
|
1/8
|
Therefore, RGH= 0.125
Cousin marriages will again increase the number of paths as they did with direct
relationships.

Combined Relationships
It is possible, if there has been any inbreeding such as a cousin marriage, for two people to be both directly and collaterally related. Below we see the calculation of the Coefficient of Inbreeding for cousin marriage. B is directly related to I (grandmother). But B is also collaterally related to I (great aunt). The second relationship results from the fact that B and I have common ancestors in C and D:

Other strategy first cousins:
Take, for example, the mating of first cousins who, by definition, share
a set of grandparents. For any particular gene in the male, the chance
that his female first cousin inherited the same gene from the same source
is 1/8. [AB x (BC or BD)=.5^3]. Further, for any gene the man passes to his child,
the chance is 1/8 that the woman has the same gene and ½ that she transmits
that gene to the child so 1/8 x ½ = 1/16. Thus, a first-cousin marriage has a
coefficient of inbreeding F =1/16= 6.25%.
http://www.genetic-genealogy.co.uk/supp/DirectRelationships.html http://www.genetic-genealogy.co.uk/Toc115570136.html
http://www.genetic-genealogy.co.uk/Toc115570135.html
CALCULATION OF THE COEFFICIENT OF RELATIONSHIP
Common Ancestors Defined
Common ancestors are usually only shared by collateral relatives, since with direct relationships one individual is the ancestor of the other. Exceptions are shown here.
In its literal sense a 'common ancestor' is any person who passes genes directly to two or more individuals. However, for the purposes of calculating the coefficient of relationship (R) and the coefficient of inbreeding (F), it has a more restricted definition:
In its narrow sense a common ancestor is any preceding individual in the pedigree who passes genes directly to two or more persons through different independent routes or paths. In the following diagram there are three individuals who satisfy this definition: i.e. B, H and I, who are common ancestors of J and K. (Note that parents are common ancestors in this context).
The coefficient of relationship ("r") between two individuals J and K is obtained by a summation of coefficients calculated for every line by which they [together] are connected to their common ancestors. Each such line connects the two individuals via a common ancestor, passing through no individual which is not a common ancestor more than once.
The coefficient of relationship ("r") between two individuals J and K is obtained by a summation of coefficients calculated for every line by which they [together] are connected to their common ancestors. Each such line connects the two individuals via a common ancestor, passing through no individual which is not a common ancestor more than once.

B passes genes directly to J and K through the following independent routes:
(B E H J) and (B F I K)
(B E H K) and (B F I J)
(B E H J) and (B F I K)
(B E H K) and (B F I J)

Similarly H and I pass genes to J and K as follows:
(H J) and (H K)
(I J) and (I K)

(H J) and (H K)
(I J) and (I K)

It can be seen that although A, C, D, E, F and G all pass genes directly to both J and K, they do not do so by different independent routes. e.g. The paths from A to J and K are:
(A E H J) and (A E H K)
(A E H J) and (A E H K)
They do not, therefore, qualify as common ancestors when calculating R . This distinction is important in finding all the relevant common ancestors in the next section. The contributions of A, C, D, E, F and G towards the degree of relationship between J and K are accounted for by the paths using H and I as common ancestors.
----------------------------
----------------------------





This work is licenced under a Creative Commons Attribution-Non Commercial-ShareAlike 4.0 International Licence.




.jpg)
.jpg)


.jpg)
.jpg)



.jpg)

.jpg)













.jpg)



.jpg)









.jpg)

.jpg)

.jpg)







{kind=link}